| 【文献翻译】密码子优化是mRNA稳定性的主要决定因素 | 您所在的位置:网站首页 › 半衰期的表示 › 【文献翻译】密码子优化是mRNA稳定性的主要决定因素 |
【文献翻译】密码子优化是mRNA稳定性的主要决定因素
|
今天给大家翻译的是2015年《CELL》期刊文章:Codon optimality is a major determinant of mRNA stability。 简要密码子的使用既在翻译水平上影响基因表达,也在mRNA衰退水平上影响基因表达,最佳和非最佳密码子之间的平衡有助于微调mRNAs水平,并最终影响蛋白质水平。 摘要信使RNA降解是基因表达的关键调控步骤。虽然已经确定了转换的主要途径,但对不同半衰期的解释仍然难以捉摸。我们发现密码子最优性是对mRNA稳定性有很大贡献的一个特征。全基因组RNA衰变分析表明,稳定的mRNA富含最佳密码子,而不稳定的mRNA则主要含有非最佳密码子。将最佳密码子替换为同义的非最佳密码子会导致mRNA的不稳定,而相反的替换会显著增加稳定性。此外,我们证明了密码子最优性影响核糖体易位,通过密码子最优性连接翻译延伸和降解过程。最后,我们表明,最佳密码子含量解释了在编码具有协调生理功能的蛋白质的mRNA中观察到的类似稳定性。这项工作表明,密码子优化作为一种机制存在,以微调mRNA水平,并最终调整蛋白质。 介绍信使RNA (mRNA)降解在调节细胞转录水平中起着关键作用,并且是调节基因表达的主要控制点。酿酒酵母中大多数mRNA的降解起始于去除3'聚腺苷酸尾(去腺苷酸化),随后切割5' 7mGppN帽(脱帽)和mRNA体在5'-3'方向的外切核酸降解。尽管被共同的降解途径靶向,但单个酵母mRNAs的周转率显著不同,半衰期从图一 从poly(A)+相对于总mRNA计算的半衰期显著不同对poly(A)+和从rpb1-1转录关闭实验制备的总RNA文库进行RNA-seq,历时60分钟。 (A)在两个文库中具有可靠半衰期的所有mRNAs都被可视化作图。颜色强度代表剩余的标准化mRNA(对于每种mRNA,时间0设置为100%)。(B)根据总mRNA测序相对于poly(A)测序计算出的每种mRNA的半衰期。差异超过2倍的数据点以红色突出显示。(C)两个图书馆的半衰期分布概况。另见表S1。 有了这些数据,我们试图识别可能决定稳定性或不稳定性的序列基序,但没有成功。跟进之前的观察,即在原本稳定的mRNA的开放阅读框中包含10个连续的稀有密码子会导致稳定性的急剧下降,我们检查了我们的转录组范围的mRNA半衰期数据,以确定ORF内的密码子含量是否会影响mRNA的稳定性。为了做到这一点,我们确定了富含任何单个密码子的mRNAs是否表现出更大或更小的稳定性。我们将信使核糖核酸定义为稳定的,如果它们的半衰期比平均值长2倍以上(~20分钟),而不稳定的,则如果它们的寿命小于平均值的一半(~5分钟)。对于每个密码子,我们计算了该密码子在mRNAs中出现的频率与mRNAs稳定性之间的相关性。将一个密码子的出现与每个mRNA的半衰期进行比较,并使用皮尔逊相关性计算产生R值(图S1E中以图形方式表示样本密码子)。我们将这一指标称为密码子出现率与mRNA稳定性相关系数(CSC)。然后将所有密码子的CSC值相互比较(图2a)。值得注意的是,观察到一些密码子优先出现在稳定的mRNAs中,而另一些密码子优先出现在不稳定的mRNAs中(总p值=1.496e-14,排列p值图2密码子组成与稳定性相关。(A)根据总RNA数据组计算,为每个密码子绘制密码子出现与mRNA稳定性相关系数(CSC)。CSC是该密码子的出现和mRNA半衰期之间相关性的R值。总p值为6.3932e-16,置换p值; 10 -4。 几十年来,大量文献假设某些密码子的翻译效率可能比其他的更高。Reis等人提出了一种衡量密码子翻译效率的方法,并将其命名为tRNA适应性指数(tAI)。这一指标旨在反映核糖体利用tRNA的效率。术语密码子优化性已被引入以试图定义翻译装置对密码子的差异识别。Frydman及其同事一般将任何阿泰高于0.47的密码子定义为最优,将任何阿泰低于0.47的密码子定义为非最优(图2B)。他们对密码子的最终命名也考虑了基因组中某些密码子的过度表达和表达不足,称为密码子偏好性(图2B;标有*)。因此,密码子优化在某种程度上反映在基因组密码子使用中(图S1A);然而,常见的密码子可以是最佳或非最佳的,而不常见的密码子也可以是最佳或非最佳的(图S1B)。引人注目的是,与稳定或不稳定mRNAs相关的密码子分别几乎完美地反映了它们的最优或非最优分配(图2C)。  图2密码子组成与稳定性相关。 图2密码子组成与稳定性相关。(B)以与(A)相同的顺序绘制的每个密码子的tRNA适应性指数值。Pechmann和Frydman (2013)中定义的密码子优化是用颜色编码的,绿色代表最佳密码子,红色代表非最佳密码子。根据其中讨论的附加标准,标有星号(*)的密码子被称为最优或非最优。 (C)如在(A)中为每个密码子绘制的密码子出现与mRNA稳定性相关系数(CSC ),但是在(B)中呈现的优化信息通过颜色编码添加。绿色代表最佳密码子,红色代表非最佳密码子。 我们的CSC指标和tAI之间的直接比较揭示了这些值之间非常好的总体一致性(图2D;R = 0.753,p值= 2.583e-12,置换p值< 10 -4)。  图2密码子组成与稳定性相关。(D)当ORF被认为是框内时,tRNA自适应索引值与CSC的关系图。绿色表示最佳密码子,红色表示非最佳密码子(R=.7255,p值为p值=2.075e-09,置换p值;10−4)。 图2密码子组成与稳定性相关。(D)当ORF被认为是框内时,tRNA自适应索引值与CSC的关系图。绿色表示最佳密码子,红色表示非最佳密码子(R=.7255,p值为p值=2.075e-09,置换p值;10−4)。重要的是,最佳密码子含量和mRNA半衰期之间的关系与用于确定半衰期的方法无关。我们使用Miller等人获得的mRNA衰变率,重复了我们对密码子使用与mRNA半衰期的分析。与我们自己的相比,这些数据是通过使用代谢标记的稳态方法计算获得的,代谢标记对细胞的干扰最小,与我们的方法完全不同。两个数据集都显示了最佳密码子含量和mRNA衰减率之间相似且惊人的相关性(图S1C和D)。 为了确定密码子优化相关性是否可能掩盖了实际上可能决定mRNA半衰期的其他特征(例如序列含量、GC百分比或二级结构),我们在通过计算引入+1和+2移码后重新分析了我们的数据。在这些移码ORF的分析中,密码子含量和稳定性之间的相关性完全消失,从而消除了其他具有决定性的变量(图2E;R=−0.127,p值=0.3303,排列p值=0.8847和图2F;R=−0.288,p值=0.0242,排列p价值=0.0012)。  图2密码子组成与稳定性相关。(E)当ORF被一个核苷酸移码时,tRNA适应性指数与CSC的关系图。绿色表示最佳密码子,红色表示非最佳密码子。(F)当ORF被两个核苷酸移码时,tRNA适应性指数与CSC的关系图。绿色表示最佳密码子,红色表示非最佳密码子。另请参见图S1。稳定和不稳定的mRNAs表现出不同的最佳密码子含量 图2密码子组成与稳定性相关。(E)当ORF被一个核苷酸移码时,tRNA适应性指数与CSC的关系图。绿色表示最佳密码子,红色表示非最佳密码子。(F)当ORF被两个核苷酸移码时,tRNA适应性指数与CSC的关系图。绿色表示最佳密码子,红色表示非最佳密码子。另请参见图S1。稳定和不稳定的mRNAs表现出不同的最佳密码子含量如上所示,我们的总mRNA稳定性数据的计算分析揭示了密码子出现和mRNA半衰期之间的关系。这些数据表明,或者是特定的密码子改变了稳定性,或者是mRNA中的整体密码子含量共同影响了稳定性。为了在个体转录物水平上评估最佳密码子含量和衰变率之间的关系,对所有个体转录物的密码子使用进行作图(图S2)。聚类分析显示,不同的mRNAs偏向于使用不同类型的密码子。总的结果并不令人惊讶,因为密码子偏好性已经得到了很好的研究;然而密码子使用的模式表明某些类别的mRNAs主要使用最佳或非最佳密码子(图3A和B;代表过多的密码子用黄色表示,代表不足的密码子用蓝色表示),并且这种用法与总体转录物稳定性相关(图3C)。对几个稳定的mRNAs的更仔细的检查揭示了这些转录物不富含任何特定的密码子,但是绝大多数比例(> 80%)的密码子属于最佳类别(图3D)。相比之下,发现单个不稳定的mRNAs在非最佳密码子中富集(60%或更多)(图3E)。这些分析表明,在这组mRNAs中,稳定的mRNA偏向于包含占优势的最适密码子,而不稳定的mRNA富含非最适密码子,尽管特定的密码子同一性在各个转录本之间有所不同。  图3.(A)一类具有相似密码子使用的相对稳定的mRNAs的热图。每一列代表单个密码子的使用,每一行代表一个mRNA.。黄色表示该密码子的使用率高于平均水平,蓝色表示低于平均使用率。有关完整的热图,请参见图S2。(B)与(A)中一样,但显示了相对不稳定的mRNAs类别。 图3.(A)一类具有相似密码子使用的相对稳定的mRNAs的热图。每一列代表单个密码子的使用,每一行代表一个mRNA.。黄色表示该密码子的使用率高于平均水平,蓝色表示低于平均使用率。有关完整的热图,请参见图S2。(B)与(A)中一样,但显示了相对不稳定的mRNAs类别。 图3.多个密码子富含在稳定和不稳定的mRNA类中。(C)显示(A)和(B)所示信使核糖核酸类别的半衰期分布的点图。 图3.多个密码子富含在稳定和不稳定的mRNA类中。(C)显示(A)和(B)所示信使核糖核酸类别的半衰期分布的点图。 图3(D)选定的稳定信使核糖核酸的密码子最优化图。基因被分解,并被绘制成单独的密码子。密码子是以最佳顺序呈现的,而不是以它们的自然顺序呈现的。更高的条形代表更理想的密码子(y轴上的CSC)。绿色表示最佳密码子,红色表示非最佳密码子。(E)所选不稳定信使核糖核酸的密码子最优化图,如(D)所示。 图3(D)选定的稳定信使核糖核酸的密码子最优化图。基因被分解,并被绘制成单独的密码子。密码子是以最佳顺序呈现的,而不是以它们的自然顺序呈现的。更高的条形代表更理想的密码子(y轴上的CSC)。绿色表示最佳密码子,红色表示非最佳密码子。(E)所选不稳定信使核糖核酸的密码子最优化图,如(D)所示。 图3.多个密码子富含在稳定和不稳定的mRNA类中。(D,E) 图3.多个密码子富含在稳定和不稳定的mRNA类中。(D,E) 图3.多个密码子富含在稳定和不稳定的mRNA类中。(F)将信使核糖核酸半衰期划分为最优组的盒图。一半的数据属于盒装部分,而胡须代表其余的数据。数据点下降到四分位数之间距离的1.5倍以上被视为异常值。 图3.多个密码子富含在稳定和不稳定的mRNA类中。(F)将信使核糖核酸半衰期划分为最优组的盒图。一半的数据属于盒装部分,而胡须代表其余的数据。数据点下降到四分位数之间距离的1.5倍以上被视为异常值。将该分析扩展到整个转录组的水平,当用百分位数评估mRNA中最佳密码子的比例时,观察到最佳密码子含量和mRNA稳定性之间的相关性。具体来说,具有少于40%最佳密码子的mRNAs通常被发现是不稳定的,平均半衰期为5.4分钟。相反,发现具有70%或更高最佳密码子含量的mRNAs是稳定的,中位半衰期为17.8分钟(图3F)。 优化密码子含量直接影响mRNA的衰减速率为了在实验上验证计算分析中观察到的关系,我们评估了改变mRNA中最佳密码子百分比对稳定性的影响。我们通过对60个非最佳密码子中的52个进行同义最佳替换来修饰不稳定的LSM8 mRNA(半衰期= 4.65分钟)的密码子含量。类似地,我们用同义的非最佳密码子替换了稳定RPS20 mRNA(半衰期= 25.3分钟)编码区内的大多数最佳密码子(113个中的108个)。该方法确保由这些序列编码的多肽与天然形式相比没有变化。此外,选择取代以避免显著改变编码区的GC含量或引入任何预测的RNA二级结构(数据未显示)。转录抑制后这些mRNAs的Northern印迹分析显示,这两个转录物中密码子的改变导致其稳定性的显著变化。具体而言,由于非最佳密码子在其ORF中转化为同义最佳密码子,LSM8 mRNA的半衰期增加了7倍以上(半衰期=18.7分钟;图4A)。相反,在稳定的RPS20 mRNA中用非最佳密码子替换最佳密码子导致其稳定性急剧降低(10倍)(半衰期= 2.5分钟;图4B)。这些数据表明,mRNA中密码子的同一性可以强烈影响稳定性,并且最佳密码子含量对决定体内mRNA的衰减速率有显著贡献。  图4.通过改变LSM8的密码子优化(A)密码子优化图可以控制mRNAs的稳定性(如图3E),显示了一个自然非优化的mRNAs。LSM8 OPT是LSM8的同义词替代版本,旨在实现更高的最优化。右侧显示了rpb1-1关闭实验的Northern斑点,两个记者的半衰期都是如此。量化归一化为SCR1加载控件(未显示)。 图4.通过改变LSM8的密码子优化(A)密码子优化图可以控制mRNAs的稳定性(如图3E),显示了一个自然非优化的mRNAs。LSM8 OPT是LSM8的同义词替代版本,旨在实现更高的最优化。右侧显示了rpb1-1关闭实验的Northern斑点,两个记者的半衰期都是如此。量化归一化为SCR1加载控件(未显示)。(B)如(A)所述,除了自然最优的信使核糖核酸外,RPS20(如图3D所示)已被设计为比RPS20非OPT更低的最佳性。RPB1-1关闭实验的Northern斑点显示在右侧,两个消息的半衰期都是如此。量化归一化为SCR1加载控件(未显示)。 为了进一步检验最佳密码子含量和mRNA稳定性之间的关系,我们产生了两个合成的开放阅读框,它们编码相同的59个氨基酸多肽,但在每个密码子的最佳性上不同(SYN报告子;图S3A、B和C)。我们将合成的ORF引入带有MFA2的5'和3'UTR的报道基因中,MFA2是一种被充分研究过的mRNA,它在细胞中被迅速降解(半衰期= 3.0分钟),这种现象部分地被其3'UTR内编码的元件介导。我们还将合成的ORF引入具有PGK1的5'和3' UTR的报道基因中,PGK11是一种特征明确且稳定的mRNA(半衰期= 25分钟)。当通过转录关闭分析测量四种报道基因mRNAs的稳定性时,发现编码最佳SYN ORF的转录物比它们携带非最佳密码子的对应物明显更稳定(约4倍)(图4C)。  (C)编码所示多肽的合成mRNA (SYN)的密码子优化图。 (C)编码所示多肽的合成mRNA (SYN)的密码子优化图。肽是人工设计的,与任何已知的蛋白质都没有相似之处。SYN opt和non-opt都是从稳定转录本(PGK1)和不稳定转录本(MFA2)插入到侧翼区域。右侧的Northern blots显示GAL关闭实验,表明SYN mRNA在MFA2和PGK1侧翼序列中具有稳定性。定量归一化为未显示的SCR1加载控制。另请参见图S3。 重要的是,最佳和非最佳编码的SYN报道基因mRNA的降解被确定为通过用于降解酵母中大多数内源mRNA的去腺苷化依赖性脱帽途径发生,而不是由已知靶向异常mRNA的三种途径中的任何一种介导(图S3G和H)。对这些mRNAs衰减的高分辨率northern分析证实,正常衰减途径中受调节的步骤去腺苷酸化和脱帽速率受到报道基因ORF内密码子组成变化的影响(图S3D、E和F)。这些数据表明,最佳密码子含量是mRNA稳定性的关键决定因素,影响mRNA转换过程中的去腺苷化和脱帽速率,而不依赖于5'和3' UTRs,它们可以平行作用以稳定或去稳定mRNA。 优化密码子含量影响翻译效率为了评估密码子优化对体内mRNA翻译效率的影响,我们产生了三种新的报道基因,它们在最佳密码子含量上不同,但在氨基酸序列上没有差异。具体而言,我们改造了HIS3基因的ORF,使其包含所有最佳密码子(HIS3 opt)或所有非最佳密码子(HIS3 non-opt),野生型HIS3基因在43%最佳密码子处提供了一个中间点(图5A)。选择HIS3基因是因为与我们的其他同义突变构建体相比,它具有相对长的ORF (220个氨基酸),允许我们通过蔗糖密度梯度有效地监测核糖体结合(见下文)。然后,我们使用rpb1-1菌株通过转录关闭分析确定了三个HIS3构建体的mRNA衰减率。与我们之前的结果一致,观察到改变最佳密码子含量对mRNA半衰期产生显著影响(图5B)。值得注意的是,对HIS3 mRNA衰减的影响与使用的最佳密码子的百分比相匹配。最佳构建体的半衰期(半衰期> 60分钟)比野生型构建体(半衰期= 9.5分钟)长得多,野生型构建体的半衰期明显长于非最佳构建体(半衰期= 2.0分钟)。因此,我们可以通过改变最佳密码子含量,在不改变蛋白质序列或侧翼序列的情况下,在酵母中实现全范围的mRNA半衰期改变。 我们假设密码子优化会影响翻译延伸。我们用两种方法测试了这个假设。首先,我们通过western blot监测HIS3最佳构建体与HIS3非最佳构建体的蛋白质产量,然后将蛋白质表达标准化为mRNA水平,如northern blot所确定的。我们观察到非最佳构建体的蛋白质产量比最佳构建体低四倍(图5C)。其次,我们评估了HIS3 mRNA结构上的核糖体密度。使用蔗糖梯度监测核糖体密度,然后对分离的级分进行分级和northern印迹。关键的是,观察到所有三种HIS3报道基因mRNAs的核糖体占据率几乎相同(图5D)。因此,我们认为,蛋白质产量减少四倍,同时在多聚核糖体中的定位几乎相同,这表明与最佳构建体相比,非最佳构建体的核糖体易位率降低。  图5在不改变核糖体关联的情况下,最优性可以影响mRNA的翻译和稳定性。(A)具有中间半衰期的转录物HIS3的密码子最优性图,以及通过同义替换分别含有较高和较低百分比的最佳密码子的版本,HIS3 opt和HIS3 non-opt。 图5在不改变核糖体关联的情况下,最优性可以影响mRNA的翻译和稳定性。(A)具有中间半衰期的转录物HIS3的密码子最优性图,以及通过同义替换分别含有较高和较低百分比的最佳密码子的版本,HIS3 opt和HIS3 non-opt。(B)rpb1-1关闭实验的Northern印迹显示了所有三种信息的半衰期。定量归一化为SCR1;加载控制未显示。 (C)最佳和非最佳版本HIS3的稳态浓度的Northern和Western印迹。加载控制和定量如下所示。翻译效率计算为相对蛋白质水平除以相对mRNA水平,并在底部绘制。对数字数据扫描进行处理,从单个凝胶图像中去除不相关的通道。 (D)微量蔗糖密度梯度分析,以及梯度组分的northern blot分析。印迹显示了三个HIS3报告基因在梯度内的位置。每个分数的定量如下所示。 优化密码子含量影响核糖体易位为了直接确定核糖体在含有非最佳密码子和最佳密码子的mRNAs上转运是否较慢,我们监测了这两种报告基因的核糖体流失。为此,我们通过剥夺细胞葡萄糖10分钟来阻断翻译起始。葡萄糖剥夺导致翻译起始的快速抑制,因此大量多聚核糖体因流失而丢失(图6A对C)。  图6最优转录本和非最优转录本在多糖体上的保留不同。(A)蔗糖密度梯度分析的代表性A260痕量显示正态分布到RNP, 80S和多核糖体部分。(B)正常条件下,最佳和非最佳HIS3报告基因以及RPS20和LSM8 mrna在蔗糖密度梯度上的分布表明,定位主要在多核糖体部分。 图6最优转录本和非最优转录本在多糖体上的保留不同。(A)蔗糖密度梯度分析的代表性A260痕量显示正态分布到RNP, 80S和多核糖体部分。(B)正常条件下,最佳和非最佳HIS3报告基因以及RPS20和LSM8 mrna在蔗糖密度梯度上的分布表明,定位主要在多核糖体部分。(C)径流条件下蔗糖密度梯度分析的代表性A260痕量,显示多核糖体组分崩塌。 为了监测核糖体流失,我们在葡萄糖剥夺前后提取mRNA-核糖体复合物,用蔗糖梯度分离材料,收集级分,并通过northern分析监测每个级分中HIS3 mRNAs的存在。重要的是,在正常条件下,确定HIS3 opt和非opt构建体的核糖体占有是相似的(图6B);然而,在诱导核糖体流失时,大部分最佳构建体mRNA重新定位到无核糖体区域的梯度顶部,而HIS3非opt mRNA仍然主要与多聚核糖体结合(图6D)。  图6(D)在径流条件下最佳和非最佳HIS3报告基因以及RPS20和LSM8 mRNAs的分布,证明了差异重定位。 图6(D)在径流条件下最佳和非最佳HIS3报告基因以及RPS20和LSM8 mRNAs的分布,证明了差异重定位。我们将这种分析扩展到密码子优化性显著不同的两种内源性mRNA转录物,LSM8 (45%最佳密码子)和RSP20 (92%最佳密码子)。值得注意的是,内源性LSM8 mRNA在翻译起始抑制后保留在多聚核糖体上,而RPS20 mRNA有效解离。我们认为保留率的差异是由于具有高最佳密码子含量的信息上更有效的核糖体易位。因此,在多核糖体组分中保留主要带有非最佳密码子的mRNAs表明密码子的最佳性可以直接影响核糖体易位的速率。 基因表达的精确性是通过协调优化密码子含量来实现的先前对酵母中mRNA稳定性的分析表明,一些编码在相同途径中起作用或属于相同复合物一部分的蛋白质的mRNA的衰变速率是相似的。单个mRNAs的更新似乎是基于它们编码的蛋白质的生理功能和细胞需求。我们假设,最佳密码子含量的调节可能为细胞协调表达共同功能蛋白质的转录物的代谢提供机制。我们评估了蛋白质产物在共同途径和/或复合体中发挥作用的基因的密码子使用情况。我们观察到,编码糖酵解相关酶的mRNAs(n=10)具有相似的和非常高的最佳密码子比例(平均值=86%;图7A)。这些转录本在之前和我们的全基因组分析中都被确定为稳定的(中位数半衰期=43.4min)。相反,编码参与酵母细胞信息素反应的多肽的mRNAs(n=14)都是不稳定的(中位半衰期=5.6min),平均只有43%的最佳密码子(图7A)。我们的分析显示,其他成绩单的表现类似。稳定的大和小核糖体亚基蛋白mRNAs(n=70和54,中位半衰期分别为18.9min和20.2min)平均最适密码子含量分别为89%和88%,但编码线粒体中核糖体蛋白的mRNAs不稳定(n=42,中位半衰期=4.8min),与观察到的45%最适密码子含量一致。(图7A和图B)。  图7.功能相关的基因显示相似的最佳性。 图7.功能相关的基因显示相似的最佳性。(A)其蛋白质产物具有相关功能的基因组被绘制出来以显示它们的最佳性。一半的数据属于盒装部分,而胡须代表其余的数据。四分位数距离下降1.5倍以上的数据点被视为异常值。代表的基因群包括70个大核糖体亚基蛋白基因、54个小核糖体亚基蛋白基因、42个线粒体核糖体蛋白基因、14个信息素反应基因、10个糖酵解酶基因、15个小亚单位加工小体基因和12个tRNA加工基因。 (B)细分两组,以显示最佳密码子含量与组内半衰期之间的关系。细胞质核糖体和线粒体核糖体中每种蛋白质的信使核糖核酸半衰期与该信使核糖核酸的最佳密码子含量相对应。 具有类似衰减率的其他基因家族包括那些其蛋白质产物参与核糖体加工、tRNA修饰、TCA循环、RNA加工和翻译机制组件的基因(图7和未显示的数据)。这些数据提供了证据,表明表达相关功能蛋白质的转录本在最佳密码子含量和衰减率水平上是协调的,这表明这些基因可能已经进化出特定的密码子内容作为一种机制,以促进基于它们在细胞中的功能的精确同步表达。 讨论我们已经提供了几条证据表明密码子的最佳性是芽殖酵母中信使核糖核酸稳定性的主要决定因素。首先,生物信息学分析表明,最佳密码子的百分比与信使核糖核酸半衰期之间存在很强的相关性。例如,具有低于40%最佳密码子的mRNAs的中位半衰期为5.3分钟。而具有超过70%最佳密码子的mRNAs的中位半衰期为20.1min。从生物信息学中得出的结论得到了实验验证,表明将最优密码子改变为非最优不稳定的mRNA,而将非最优密码子改变为最优稳定的mRNA,否则不稳定。最重要的是,我们提供了最优和非最优密码子通过调节翻译延伸率发挥作用的证据。 几项核糖体图谱研究未能发现最佳密码子和非最佳密码子翻译中特定密码子的差异。然而,当编码相同多肽的mRNAs由最佳密码子或非最佳密码子组成时,我们观察到核糖体清除方面的显著差异。这些差异可能反映了许多小核糖体在非最佳密码子上的犹豫不决的加性效应。在核糖体图谱分析中,这种犹豫是不可察觉的。或者ORF的整体密码子组成可以设置整个ORF的统一的翻译延伸率。如果这是真的,核糖体图谱就不会检测到单个密码子的速率变化。 值得注意的是,虽然密码子含量显然是mRNA稳定性的主要决定因素,但它并不能预测所有mRNAs的半衰期。例如,几种组蛋白成分的信使核糖核酸,如HHF2和HHT1,含有85%的最佳密码子,但非常不稳定,半衰期分别为2.4和3.5分钟。这种mRNAs的半衰期可以由它们高效(或低效)启动翻译的能力和/或由5'或3' UTRs中的元件决定。每种方法都有大量的例子被描述。开放源码框架内的特征也可能解释一些异常值(例如优化和非优化密码子的分布)。 由于最佳密码子含量对翻译延长率的影响,很可能是某些因素(S)在mRNAs与核糖体接触时监控这些速率。事实上,我们之前已经表明,通过插入稀有密码子来减缓核糖体的运动会促进mRNA的降解。监测因子的主要候选是DEAD-box RNA解旋酶DHH1,它是mRNA衰变机制的一个组成部分,已被证明通过促进翻译抑制的作用作为脱帽的激活剂。翻译延伸率影响信使核糖核酸衰变的机制还需要进一步研究。 通过密码子优化实现基因表达的精确度和协调性长时间尺度和短时间尺度都为细胞内密码子最佳性的重新分配提供了重要机会。在短期内,细胞生长条件和营养可获得性的变化可能显著影响单个(或部分)带电tRNA水平。这种供应减少的结果是,由于周转增加,富含在这些tRNA解码的密码子中的mRNAs的翻译延伸速度将减慢,其水平将下降。通过这种方式,密码子最优化不仅为细胞提供了磨练mRNA水平的一般机制,也为细胞提供了感知环境条件和快速定制基因表达的全球模式的机制。 长期的遗传变化可以将同义突变引入蛋白质编码基因,这些突变不会改变编码多肽的氨基酸序列;然而,如果突变显著改变了信使核糖核酸开放阅读框架内最佳密码子的比例,这种变化将影响信使核糖核酸和蛋白质表达水平。因此,同义基因突变可以被设想为一种进化对细胞有利的信使核糖核酸稳定率的方法。我们发现,编码蛋白质的mRNAs在相似的路径上共同作用,或者是相同化学计量复合体的一部分,并且以前被观察到以类似的速度降解,编码几乎相同比例的最佳密码子(图7A)。 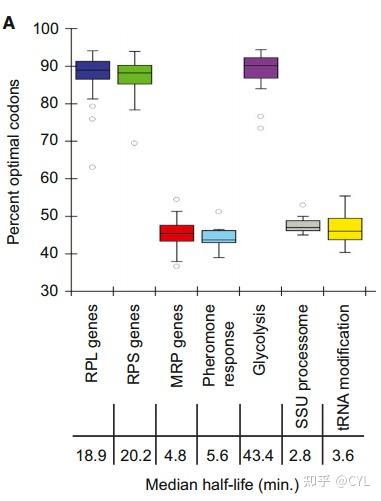 我们认为,密码子的最优化已经针对这些基因集进行了微调,作为一种优雅的机制,以确保转录后协调调节和蛋白质在细胞所需的精确水平上的简约表达。有趣的是,相似水平的最佳密码子和非最佳密码子不仅可以确保相关mRNAs的稳定性和翻译率的相似性,而且还可以确保对tRNA水平变化(例如营养可获得性、胁迫、细胞类型等)的反应协调。最近的研究表明,细胞内的tRNA浓度不是静态的,而是不断地发生变化,有时是戏剧性的。例如,大规模RNA图谱实验表明,tRNA浓度在增殖细胞和分化细胞之间差异很大。根据我们的分析,我们认为tRNA浓度的显著变化可以通过动态改变mRNA的稳定性来改变细胞内的mRNA表达谱,即使转录没有任何变化。 核糖体是主要的把关人,决定正常和异常mRNAs的下游命运最后,我们的工作表明,核糖体的共翻译mRNA监控不仅对快速降解异常mRNA很重要,而且对调节正常mRNA的降解速率也很重要。在真核生物中,mRNA的异常导致异常翻译事件,如过早终止、翻译终止缺失和核糖体停滞,这分别通过无意义介导的、Non-Stop和No-Go降解途径导致mRNA的加速周转。我们发现正常mRNA中的密码子使用也影响翻译核糖体,并对mRNA的稳定性有深远的影响。因此,核糖体作为主传感器,通过调节其延伸和/或终止过程,帮助确定所有正常和异常mRNAs的命运。使用核糖体作为传感器是蛋白质编码基因的理想选择,蛋白质编码基因在细胞中的主要功能是翻译。我们认为,mRNA稳定性的一个组成部分作为密码子组成的一个功能被构建到所有的mRNA中。翻译核糖体的延伸速率被传递给一般的降解机制,这影响去腺苷化和脱帽的速率。单独来说,一个mRNA中密码子的同一性预计对整个核糖体的解码有微小的影响;然而,在一个完整的mRNA框架内,我们发现密码子优化对翻译延伸和mRNA更新有深远的影响。因此,我们得出结论,密码子同一性代表了mRNAs的一个普遍性质,并且是它们稳定性的一个关键决定因素。 PDF原文下载链接1: 或者2: |
【本文地址】